S. Casano, G. Grassi, V. Martini i M. Michelozzi (2011)
Variations in terpene profiles of different strains of Cannabis sativa L. Acta Horticulturae 925:115-121
Els compostos causants dels efectes psicotròpics del Cànnabis, com el principal cannabinoide psicotròpic Δ9-THC (delta9-tetrahydrocannabinol), han estat àmpliament estudiats per la possible utilitat en aplicacions medicinals. La indústria farmacèutica, en canvi, està interessada en la planta com a font de matèria prima i estudia la variabilitat i sinergia entre els diversos metabòlits secundaris. Altres cannabinoides, terpenoides i flavonoides poden reduir l’ansietat, el dèficit colinèrgic i la immunosupressió provocada pel Δ9-THC, alhora que incrementar el flux sanguini cerebral, millorar l’activitat cortical, acabar amb patògens respiratoris i actuar com a antiinflamatoris (McPartland i Russo, 2001). Els terpenoides posseeixen una gran varietat de propietats biològiques, inclosos efectes quimiopreventius contra el càncer i la millora de la penetració de la pell, així com activitat antimicrobiana, antifúngica, antiviral, antihiperglicèmica, antiinflamatòria i antiparasitària (Paduch et al., 2007).
Les plantes presenten canvis bioquímics dinàmics quan són atacades per malalties i herbívors, així com en resposta a l’estrès abiòtic, la qual cosa dóna lloc a la producció induïda i l’alliberament de compostos volàtils de l’aroma beneficiosos per a la defensa directa o indirecta de la planta. En l’Arabidopsis thaliana (Huang et al., 2010) i la Medicago truncatula (Navia-Ginè et al., 2009) es produeix una variació quantitativa significativa en l’emissió del monoterpè trans-β-ocimè a conseqüència de l’atac d’insectes herbívors. Dos monoterpens que solen estar presents entre els compostos volàtils de l’aroma del Cànnabis, limonè i α-pinè, com altres monoterpens, han demostrat ser potents repel·lents d’insectes herbívors (Nerio et al., 2010), mentre que els sesquiterpens tendeixen a ser relacionats amb la ingesta per part d’animals herbívors. Potter (2009) va demostrar que, en el Cànnabis, la proporció monoterpè:sesquiterpè existent en les fulles i inflorescències és molt diferent a causa de la presència dominant de tricomes sèssils en el fullatge, i de tricomes capitats pedunculats en el material floral, amb els monoterpens més volàtils dominant en les fulles per actuar com a antiherbívors per a animals herbívors. Com que aquest és un Cànnabis d’ús farmacèutic, normalment es cultiva en instal·lacions inaccessibles per a animals herbívors, on es manté com a major problema el dels insectes herbívors, especialment els més comuns i destructius com són l’aranya roja, els trips, i la mosca blanca. Per tant, l’anàlisi de monoterpens i l’estudi de la seva variabilitat podria jugar un paper estratègic en la selecció de plantes menys susceptibles de l’atac d’aquests i altres insectes.
Els terpens són fortament heretats i poc influenciats per factors de l’entorn, per la qual cosa han estat molt utilitzats com a marcadors bioquímics en estudis quimiosistemàtics per definir espècies de plantes, procedència, clons i híbrids. S’ha constatat una gran variabilitat en el contingut de terpens en diferents soques de Cànnabis (Mediavilla i Steinemann, 1997; Novak et al., 2001; Hillig, 2004; Fischedick et al., 2010). La variabilitat en el metabolisme secundari combinada amb informació genètica ha reobert un antic debat sobre l’enfocament taxonòmic. De fet, Hilling (2005) va proposar un concepte politípic que reconeix tres espècies (Cannabis sativa, Cannabis indica i Cannabis ruderalis) i set suposats tàxons, però la majoria dels investigadors segueix recolzant l’enfocament monotípic i identifica les espècies com a Cannabis sativa L. La diferenciació de les soques en “sativa pura”, “predominança sativa”, “híbrid sativa/índica”, “predominança índica”, “índica pura” i “híbrid ruderalis” és adoptada generalment per conreadors i productors per diferenciar els diversos biotips. Aquest estudi va investigar la variabilitat en els perfils de terpens de soques de Cànnabis i va explorar la utilitat dels monoterpens en la distinció entre els biotips de “predominança sativa” i “predominança índica”.
Materials i mètodes
Es van obtenir diferents soques amb un perfil Δ9-THC de conreadors d’empreses privades. La designació de soques com a biotips de “predominança sativa” o “predominança índica” es va basar en l’historial genètic aportat pels conreadors. La designació de biotips d’“índica pura” o “sativa pura” no es va posar en pràctica per la falta d’informació sobre aquestes soques. Cada soca consistia en un paquet comercial amb, aproximadament, deu llavors viables. Es van realitzar avaluacions preliminars sobre historial genètic aportat plantant aquestes soques durant la temporada de primavera-estiu en un hivernacle situat a CRA-CIN (Rovigo). Al principi de l’etapa de floració es van eliminar les plantes estaminades, mentre que les plantes pistil·lades van ser tractades amb una solució de tiosulfat de plata per induir artificialment la producció d’inflorescències estaminades. Es va produir l’autopol·linització de totes les plantes pistil·lades mitjançant l’aïllament de cadascuna amb bosses blanques de paper. Només 16 plantes pistil·lades derivades de 16 soques diferents van ser seleccionades finalment. 8 plantes (identificades com a 5, 6, 7, 8, 9, 10, 11 i 12) van ser derivades de soques de “predominança sativa” i unes altres 8 (identificades com a 2, 3, 4, 13, 14, 15, 16 i 17) de soques de “predominança índica”.
Les progènies de 16 plantes es van conrear en interior a CRA-CIN (Rovigo). 99 plantes en total (de 3 a 7 plantes per cada soca) es van desenvolupar sota llums de sodi a alta pressió 600 W/m2 (Philips Son-T). El fotoperíode es va mantenir en 18 hores de llum durant les primeres 4 setmanes de cultiu i després es va reduir a 12 hores fins el moment de la collita. La temperatura i la humitat relativa de l’aire van ser mantingudes, respectivament, a 25±3 °C i 50-70%. Les plantes van ser conreades individualment en tests d’1,5 l, en una fibra de coco finíssima (CANNA B.V.), i van ser fertirrigades cada dia mitjançant un sistema d’irrigació automàtic, amb la dosi adequada en funció de les necessitats. La solució de nutrients utilitzada (EC=1,7) es va obtenir mesclant parts iguals de Coco A i B (CANNA B.V.) amb aigua de l’aixeta, i el nivell de pH es va ajustar a 5.5. La fertirrigació va ser interrompuda 2 setmanes abans de la collita i els tests van ser esbandits amb aigua de l’aixeta amb un pH=5.5.
La collita de les primeres soques (“predominança índica”) es va produir després de 105 dies de la sembra, mentre que la collita de les més tardanes (“predominança sativa”) no es va produir fins als 133 dies. Es van mostrejar teixits frescos d’inflorescències de les plantes durant la collita per a l’anàlisi de terpenoides. El material de mostra (80 mg teixits frescos d’inflorescències) va ser introduït en nitrogen líquid, extret en 4 ml de n-pentà i, posteriorment, 1 ml de l’extracte va ser transferit a vials de GC. La composició de terpens va ser analitzada per GC/FID. Es van detectar 28 compostos en total, 15 dels quals completament identificats i 13 romanen encara sense identificar (desc.). Els terpenoides van ser identificats després de fer correspondre els seus temps de retenció amb els de compostos purs sota les mateixes condicions. En funció dels temps de retenció, els màxims van ser identificats com a α-pinè, desc. 1, desc. 2, camfè, β-pinè, sabinè, Δ-3-carè, α-fel·landrè, β-mircè, α-terpinè, limonè, 1,8 cineol, γ-terpinè, cis-β-ocimeno, trans-β-ocimè, α-terpinolè, desc. 3, desc. 4, β-cariofilè, desc. 5, desc. 6, desc. 7, desc. 8, desc. 9, desc. 10, desc. 11, desc. 12 i desc. 13. La majoria dels terpens identificats van ser monoterpens, amb l’excepció d’un sesquiterpè, el β-cariofilè.
El contingut relatiu de cada monoterpè va ser expressat com a percentatge del total de monoterpens, mentre que cada sesquiterpè va ser calculat com a percentatge del total de monoterpens i sesquiterpens. La informació no va ser tractada de la manera més habitual (Kolmogorov-Smirnov One-Sample Test), sinó que va ser analitzada per l’ANOVA no paramètric de Kruskal-Wallis, seguit del Test U de Mann-Whitney per tal d’obtenir múltiples comparatives. Les diferències es van definir com a significatives a un nivell del 5%. Es van realitzar anàlisis estadístiques amb el programa SYSTAT 12.0 (Systat Software Inc., EUA).
Discussió dels resultats
El contingut relatiu de terpenoides és fortament heretat mentre que la producció total per pes del teixit està més subjecta a factors de l’entorn. L’expressió de la composició basada en un teixit (mg/g) és usada per al control de qualitat i l’estandardització de les varietats de Cànnabis conreades, així com per a estudis quimiosistemàtics (Fischedick et al., 2010), però el contingut relatiu (%) de terpenoides és més utilitzat per a estudis quimiosistemàtics.
La mitjana de continguts relatius de compostos dominants detectats en els compostos volàtils de l’aroma va ser: β-mircè (46,1±2,6%), α-pinè (14,0±1,5%), α-terpinolè (10,2±1,8%), limonè (7,3±1,3%), trans-β-ocimè (6,6±0,7%), β-pinè (6,1±0,4%), α-terpinè (3,6±1,0%), β-cariofilè (1,2±0,2%), 1,8 cineol (1,1±0,2%), α-fel·landrè (0,7±0,1%) i Δ-3-carè (0,6±0,1%). La mitjana de continguts relatius de camfè, desc. 1, cis-β-ocimè, desc. 5, desc. 8, desc. 7, desc. 13, sabinè, γ-terpinè, desc. 3, desc. 4, desc. 6, desc. 10, desc. 2, desc. 9, desc. 11 i desc. 12 va ser inferior al 0,5%.
Els resultats de l’ANOVA de Kruskal-Wallis ANOVA entre diferents soques (d.f.=15, N=99) van mostrar canvis significatius en els continguts relatius de tots els compostos: α-pinè (X2=71,6, P<0,001), desc. 1 (X2=71,5, P<0,001), desc. 2 (X2=43,6, P<0,001), camfè (X2=67,2, P<0,001), β-pinè (X2=53,2, P<0,001), sabinè (X2=72,5, P<0,001), Δ-3-carè (X2=69,4, P<0,001), α-fel·landrè (X2=59,6, P<0,001), β-mircè (X2=47,7, P<0,001), α-terpinè (X2=36,3, P<0,01), limonè (X2=77,1, P<0,001), 1,8 cineol (X2=67,5, P<0,001), γ-terpinè (X2=30,9, P<0,01), cis-β-ocimè (X2=79,5, P<0,001), trans-β-ocimè (X2=82,1, P<0,001), α-terpinolè (X2=78,7, P<0,001), desc. 3 (X2=37,6, P<0,001), desc. 4 (X2=33,7, P<0,01), β-cariofilè (X2=55,7, P<0,001), desc. 5 (X2=65,6, P<0,001), desc. 6 (X2=74,4, P<0,001), desc. 7 (X2=50,1, P<0,001), desc. 8 (X2=64,7, P<0,001), desc. 9 (X2=63,2, P<0,001), desc. 10 (X2=61,1, P<0,001), desc. 11 (X2=80,1, P<0,001), desc. 12 (X2=61,8, P<0,001) i desc. 13 (X2=52,8, P<0,001).
El β-mircè va ser detectat en un percentatge alt en totes les soques, i va ser la soca 17 la de major contingut relatiu (80,1±7,3%) i la 8 la de menor (16,1±3,4%) (Taula 1). El β-mircè va ser el terpè dominant en gairebé totes les soques a excepció de les soques 6, 7, 8 i 12. L’α-terpinolè va ser detectat en un percentatge alt de soques de “predominança sativa” (7, 8, 9, 10 i 12), amb les soques 7 i 8 amb aquest com a terpè dominant (41,8±7,2% i 37,3±3,5% respectivament), mentre que no va aparèixer o va aparèixer vagament en soques de “predominança índica” i en algunes de “predominança sativa” (5, 6 i 11). L’α-pinè i el β-pinè van ser detectats en totes les soques i els continguts relatius van ser inferiors al 10%. L’α-pinè va aparèixer en continguts relatius majors (fins a un 10%) en algunes soques (3, 6, 8, 11, 12, 14, 15 i 16), i es va mostrar com el terpè dominant en les soques 6 i 12 (46,3±5,7% i 24,2±15,6% respectivament). El β-pinè va ser detectat en continguts relatius superiors (fins a un 10%) en les soques 3 (12,6±1,6%) i 6 (13,2±0,8%). El limonè es va manifestar en percentatges inferiors o va aparèixer lleument en algunes soques “predominança índica” (3, 14, 15 i 16) i en algunes de “predominança sativa”, mentre que va ser localitzat en percentatges molt superiors (fins a un 10%) en altres soques de “predominança índica” (2, 4, 13 i 17), les quals van mostrar el limonè com a segon terpenoide més abundant. El Trans-β-ocimè no va ser detectat o ho va ser en percentatges baixos en una soca de “predominança sativa” (6) i en soques de “predominança índica”, mentre que va aparèixer en percentatges molt superiors en altres soques de “predominança sativa” (5, 7, 8, 9, 10, 11 i 12) (fins a un 5%), i en les soques 5 i 11 va ser el terpenoide més abundant (18,7±1,9% i 16,8±2,2% respectivament). L’α-terpinè va ser detectat lleument o en percentatges baixos en la majoria de les soques, però la soca 4 va destacar per tenir-ne un contingut relatiu molt superior (18,0±8,0%). El sesquiterpè β-cariofilè va ser detectat en totes les soques i el contingut relatiu va aparèixer principalment en percentatges inferiors al 2%, tot i que va mostrar continguts relatius en algunes soques (2, 9, 13 i 17) de fins al 2%. L’1,8 cineol va aparèixer en percentatges baixos (fins a un 2%) en algunes soques de “predominança sativa” (7, 8, 9, 10 i 12), mentre que va ser detectat lleument o en percentatges baixos en soques de “predominança índica” i en algunes altres de “predominança sativa” (5, 6 i 11). Tant el Δ-3-carè com l’α-felandrè es van mostrar en percentatges baixos (fins a un 1%) en algunes soques de “predominança sativa” (7, 8, 9, 10 i 12), mentre que no van aparèixer en soques de “predominança índica” ni en algunes de “predominança sativa” (5, 6 i 11).
El Test U de Mann-Whitney realitzat entre soques de “predominança sativa” i “predominança índica” (d.f.=1, N=99) va revelar canvis significatius en continguts relatius de diferents composts, a excepció de l’α-pinè, desc. 2, β-pinè, α-terpinè, γ-terpinè, β-cariofilè, desc. 7, desc. 12 i desc. 13 (Fig. 1). Els continguts relatius de camfè (X2=22,7, P<0,001), β-mircè (X2=23,1, P<0,001), limonè (X2=27,8, P<0,001), desc. 3 (X2=15,4, P<0,001), desc. 6 (X2=29,9, P<0,001) i desc. 11 (X2=42,3, P<0,001) van ser significativament superiors en les soques de “predominança índica” que en les de “predominança sativa” (Fig. 1). Les plantes provinents de les soques de “predominança sativa” van mostrar proporcions notablement superiors de desc. 1 (X2=33,4, P<0,001), sabinè (X2=24,9, P<0,001), Δ-3-carè (X2=39,6, P<0,001), α-fel·landrè (X2=31,97, P<0,001), 1,8 cineol (X2=19,2, P<0,001), cis-β-ocimè (X2=48,6, P<0,001), trans-β-ocimè (X2=52,6, P<0,001), α-terpinolè (X2=13,2, P<0,001), desc. 4 (X2=15,3, P<0,001), desc. 5 (X2=29,6, P<0,001), desc. 8 (X2=24,3, P<0,001), desc. 9 (X2=7,5, P<0,01) i desc. 10 (X2=9,5, P<0,01) que les de les trobades en les provinents de las soques de “predominança índica” (Fig. 1).
Tot i que Hillig (2004) va afirmar que les diferències de terpenoides en el Cànnabis són d’utilitat limitada per al criteri taxonòmic quant a espècies, amb els sesquiterpens generalment més útils que els monoterpens, hem trobat que diversos monoterpens poden servir com a potents eines per diferenciar entre biotips de “predominança sativa” i “predominança índica” (Taula 1 i Fig. 1). Les nostres conclusions es recolzen també en resultats recents obtinguts per Fischedick et al. (2010), que mostren que els monoterpens poden ajudar a diferenciar entre varietats amb sesquiterpens i nivells de cannabinoides semblants.
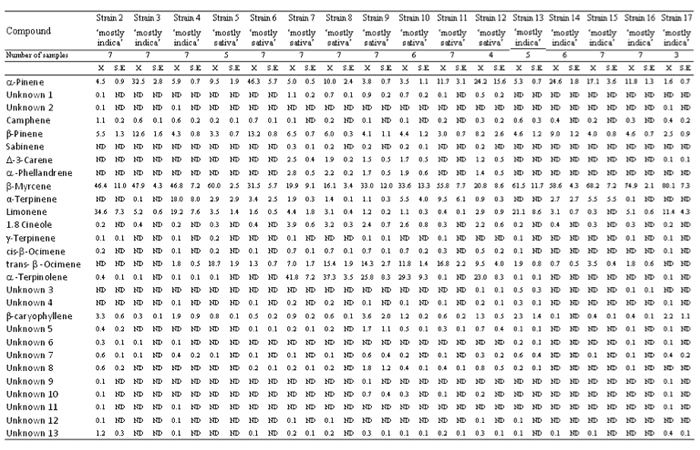
Taula 1. Perfils de terpens de diferents soques de “predominança índica” i “predominança sativa” de Cannabis sativa L.
ND = no detectat
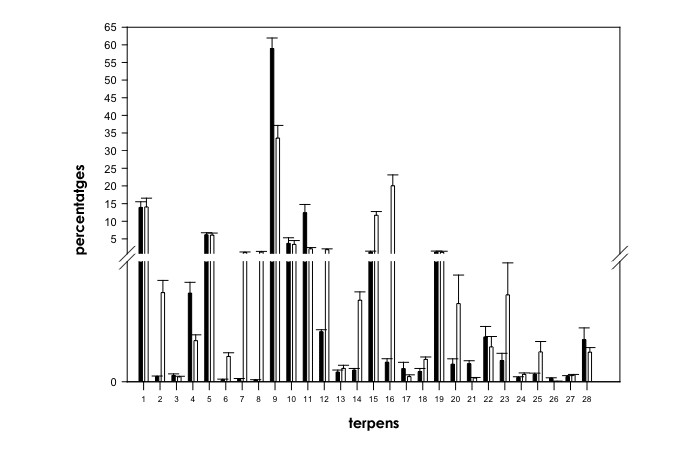
Fig. 1. Comparativa de perfils de terpens en soques de Cannabis sativa L. de “predominança índica” (histogrames negres) i “predominança sativa” (histogrames blancs). El tall en l’eix Y se situa en 0,7-0,8. Els números en l’eix X fan referència als compostos individuals següents: 1=α-pinè, 2=desc. 1, 3=desc. 2, 4=camfè, 5=β-pinè, 6=sabinè, 7=Δ-3-carè, 8=α-fel·landrè, 9=β-mircè, 10=α-terpinè, 11=limonè, 12=1,8 cineol, 13=γ-terpinè, 14=cis-β-ocimè, 15=trans-β-ocimè, 16=α-terpinolè, 17=desc. 3, 18=desc. 4, 19=β-cariofilè, 20=desc. 5, 21=desc. 6, 22=desc. 7, 23=desc. 8, 24=desc. 9, 25=desc. 10, 26=desc. 11, 27=desc. 12 i 28=desc. 13.
Conclusions
Les principals diferències entre els perfils de terpens de les soques avaluades dels dos biotips principals van ser que les soques de “predominança índica” es caracteritzaven per la dominància de β-mircè, present en continguts relatius alts, amb el limonè o l’α-pinè com els segons terpenoides més abundants, mentre que les soques de “predominança sativa” es caracteritzaven per tenir perfils de terpens més complexos, amb algunes soques manifestant l’α-terpinolè o l’α-pinè com a terpenoides dominants, i algunes soques amb el β-mircè com a terpenoide dominant i l’α-terpinolè o el trans-β-ocimè com a terpenoides secundaris més abundants.
Aquesta variabilitat en la composició de terpens pot ser una eina potent per a la qualificació de biotips de Cànnabis, així com una garantia per a investigadors futurs a l’hora d’estimar-ne el valor medicinal, i, alhora, en la selecció de quimiotips menys susceptibles de l’atac d’hervíbors i malalties, fet que constata la necessitat d’una investigació més profunda sobre la variabilitat de monoterpens i sesquiterpens. El cultiu de plantes amb terpenoides específics és un tema d’investigació fascinant; de fet, les variades activitats biològiques d’aquests compostos fan de l’anàlisi dels terpenoides una eina valuosa per a la millora d’un nombre considerable d’atributs en varietats de Cànnabis farmacèutiques i industrials.
L’anàlisi de terpenoides, combinat amb el de cannabinoides i flavonoides, és essencial per a l’empremta metabòlica de varietats de conreu farmacèutiques. Les varietats farmacèutiques dels dos biotips principals poden mostrar propietats medicinals inconfusibles a causa de diferències significatives en els continguts relatius de terpenoides, per la qual cosa la sinergia entre els diversos metabòlits secundaris ha de ser investigada en més profunditat, per aclarir el fitocomplex del Cànnabis i permetre així la selecció de quimiotips amb efectes medicinals determinats.
Agraïments
Agraïm a Phytoplant Research S.L. el recolzament financer.
Bibliografia esmentada
- Fischedick, J.T., Hazekamp, A., Erkelens, T., Choi, Y.H. and Verpoorte, R. 2010. Metabolic fingerprinting of Cannabis sativa L., cannabinoids and terpenoids for chemotaxonomic and drug standardization purposes. Phytochemistry 71:2058-2073.
- Hillig, K.W. 2004. A chemotaxonomic analysis of terpenoid variation in Cannabis. Biochemical Systematics and Ecology 32:875-891.
- Hillig, K.W. 2005. Genetic evidence for speciation in Cannabis. Genetic Resources and Crop Evolution 52:161-180.
- Huang, M., Abel, C., Sohrabi, R., Petri, J., Haupt, I., Cosimano, J., Gershenzon, J. and Tholl, D. 2010. Variation of herbivore-induced volatile terpenes among Arabidopsis ecotypes depends on allelic differences and subcellular targeting of two terpene synthases, TPS02 and TPS031. Plant Physiology 153:1293-1310.
- McPartland, J.M. and Russo, E.B. 2001. Cannabis and Cannabis extracts: greater than the sum of their parts? Journal of Cannabis Therapeutics 1:103-132.
- Mediavilla, V. and Steinemann, S. 1997. Essential oil of Cannabis sativa L. strains. Journal of the International Hemp Association 4:80-82.
- Navia-Ginéa, W.G., Yuanb, J.S., Mauromoustakosd, A., Murphye, J.B., Chenb, F. and Kortha, K.L. 2009. Medicago truncatula (E)-β-ocimeno synthase is induced by insect herbivory with corresponding increases in emission of volatile ocimeno. Plant Physiology and Biochemistry 47:416-425.
- Nerio, L.S., Olivero-Verbel, J. and Stashenko, E. 2010. Repellent activity of essential oils: a review. Bioresource Technology 101:372-378.
- Novak, J., Zitterl-Eglseer, K., Deans, S.G. and Franz, C.M. 2001. Essential oils of different cultivars of Cannabis sativa L. and their antimicrobial activity. Flavour and Fragrance Journal 16:259-262.
- Paduch, R., Kandefer-Szerszeń, M., Trytek, M. and Fiedurek, J. 2007. Terpenes: substances useful in human healthcare. Archivum Immunologiae et Therapiae Experimentalis 55:315-327.
- Potter, D. 2009. The propagation, characterisation and optimisation of Cannabis sativa L. as a phytopharmaceutical. Ph.D. Thesis in Pharmaceutical Sciences. Department of Pharmaceutical Science Research, King’s College, London.